If it could be demonstrated that any complex organ existed which could not possibly have been formed by numerous successive slight modifications, my theory would absolutely break down (Darwin, 1859, p. 219).
... The number of intermediate varieties, which have formerly existed on the earth, (must) be truly enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps, is the most obvious and gravest objection which can be urged against my theory (Darwin, 1859, p. 292).
Well, we are now about 120 years after Darwin, and knowledge of the fossil record has been greatly expanded ... ironically, we have even fewer examples of evolutionary transition than we had in Darwin's time. By this I mean that some of the classic cases of darwinian change in the fossil record, such as the evolution of the horse in North America, have had to be discarded or modified as a result of more detailed information ... (Raup, 1979).
The history of most fossil species include two features particularly inconsistent with gradualism:1) Stasis - most species exhibit no directional change during their tenure on earth. They appear in the fossil record looking much the same as when they disappear; morphological change is usually limited and directionless; 2) Sudden appearance - in any local area, a species does not arise gradually by the steady transformation of its ancestors; it appears all at once and 'fully formed' (Gould, 1977).
"is effectively dead, despite its persistence as textbook orthodoxy" (Gould, 1980).
This is true of all thirty-two orders of mammals...The earliest and most primitive known members of every order already have the basic ordinal characters, and in no case is an approximately continuous sequence from one order to another known. In most cases the break is so sharp and the gap so large that the origin of the order is speculative and much disputed...This regular absence of transitional forms is not confined to mammals, but is an almost universal phenomenon, as has long been noted by paleontologists. It is true of almost all classes of animals, both vertebrate and invertebrate...it is true of the classes, and of the major animal phyla, and it is apparently also true of analogous categories of plants (Simpson, 1944).
Taxa recognized as orders during the (Precambrian-Cambrian) transition chiefly appear without connection to an ancestral clade via a fossil intermediate. This situation is in fact true of most invertebrate orders during the remaining Phanerozoic as well. There are no chains of taxa leading gradually from an ancestral condition to the new ordinal body type. Orders thus appear as rather distinctive subdivisions of classes rather than as being segments in some sort of morphological continuum (Valentine, Awramik, Signor, and Sadler, 1991).
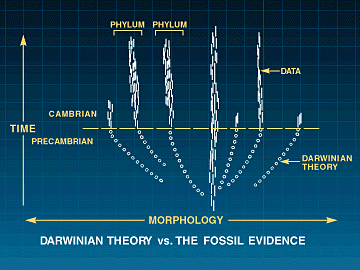
FIGURE 1: Darwinian Theory vs. the Fossil Record
The success of Darwinism was accompanied by a decline in scientific integrity. This is already evident in the reckless statements of Haeckel and in the shifty, devious and histrionic argumentation of T. H. Huxley...To establish the continuity required by the theory, historical arguments are invoked even though historical evidence is lacking. Thus are engendered those fragile towers of hypotheses based on hypotheses, where fact and fiction intermingle in an inextricable confusion (Thompson, 1956).
"Stasis is data" (Gould, 1991). Scientists cannot afford to lose sight of this abundant historical evidence. Gould recently described the importance of understanding stasis in these terms:
...we must understand that nothing happens most of the time -- and we don't because our stories don't admit this theme -- if we hope to grasp the dynamics of evolutionary change. (This sentence may sound contradictory, but it isn't. To know the reasons for infrequent change, one must understand the ordinary rules of stability.) The Burgess Shale teaches us that, for the history of basic anatomical designs, almost everything happened in the geological moment just before, and almost nothing in more than 500 million years since (Gould, 1988).
It is probably only the stasis on the level of higher taxa which is both valid and differs qualitatively from the other levels of stasis. Only higher taxa lack demonstrable evidence of change ... Higher taxon-level stasis could conceivably be the result of what might be called Baraminic Stasis -- the permanent constraint of organisms under natural conditions to stay within the bounds of their baramin (Wise, 1991).
- Title Page
- Conflicts Between Darwin and Geological Succession
- On The Origin of Stasis by Means of Natural Selection
- Facts, Fossils, and Philosophy
- References